
Big-picture breakdown
The CODES paper codes.pdf is a broad “coherence-first” framework. Its core move is to replace probability/entropy language with phase alignment language. In the paper’s vocabulary, a system is healthy/lawful/stable when its active signals remain phase-aligned above a threshold, and it fails when drift exceeds a bound. The paper names the main score PAS: a Phase Alignment Score. In biology, that becomes PAS_bio, and biological failure becomes ΔPAS_bio_zeta drift rather than merely “toxicity,” “noise,” or “entropy.”
The simplest translation is:
CODES = formal coherence language.
ceLLM/RF Safe = proposed cellular mechanism for how modern RF/ELF noise may degrade that coherence.
Entropic waste = coherence-degrading signal load.
Bioelectrical dissonance = the biological state after PAS_bio falls or ΔPAS_bio_zeta rises beyond the system’s ability to self-correct.
What CODES is claiming
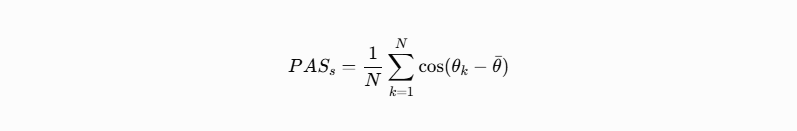
The paper’s main thesis is that reality, intelligence, biology, and emergence are not primarily stochastic. They are described as resonance systems governed by legality gates: PAS_s, ΔPAS_zeta, PAS_LOCK, TEMPOLOCK, GLYPHLOCK, AURA_OUT, and the biological analogues PAS_bio, ELF_BIO, CHIRAL_GATE, BIO_LOCK, and SOMA_OUT. In the biological section, VESSELSEED is framed as a carbon bio-coherence substrate where biological emission is permitted only when PAS_bio is high, drift is bounded, timing is lawful, and chirality is intact.
That matters because CODES does not treat “entropy” as just thermodynamic disorder. It reframes entropy as coherence loss or failed legality. In Section 15.3, the paper explicitly defines resonant entropy as a coherence deficit, and says classical entropy reappears when coherence collapses. In Section 33.6, it goes further and says entropy is the perception/signature of illegality: a system has exited lawful emission space when PAS drops below threshold or ΔPAS_zeta exceeds drift bounds.
So the CODES paper gives you a formal vocabulary for what your ceLLM/RF Safe language calls low-fidelity biology.
How this relates to entropic waste
In ceLLM terms, entropic waste is not just “energy deposited into tissue.” It is non-useful, mistimed, waveform-structured interference injected into the cell’s signaling substrate. The RF Safe/ceLLM documents describe pulsed EMF/RF as entropic waste because it may degrade the geometric fidelity of cellular intelligence, especially where bioelectric gradients, Ca²⁺ signaling, mitochondrial redox, radical-pair chemistry, and chromatin topology are being used as an information-processing layer.
CODES would restate that like this:
Entropic waste = external field perturbation that lowers PAS_bio or increases ΔPAS_bio_zeta.
Formally:
RF_state(t) → phase perturbation θ_k(t) → PAS_bio↓ / ΔPAS_bio_zeta↑ → ELF_BIO correction or BIO_LOCK failure
In other words, the waste is not merely heat. It is phase-disordering load. A clean biological signal has timing, amplitude, phase, rhythm, and coherence. A waste signal may have energy but no biological legality. It forces the system to spend correction capacity restoring order.
That fits the CODES definition almost exactly: entropy is not fundamental randomness; entropy is what the system experiences when lawful recurrence fails.
How this relates to bioelectrical dissonance
Bioelectrical dissonance, as described in the ceLLM/RF Safe material, is the state where the cell’s predictive/inference layer receives corrupted bioelectric inputs. The documents describe cells as closed-loop inference systems using bioelectric vectors, calcium timing, redox state, UPE/biophoton patterns, and DNA/chromatin topology as a kind of latent biological state space. EMF exposure is framed as “cat-picture noise” or entropic waste injected into that latent system.
CODES gives this a direct formalization:
Bioelectrical dissonance = PAS_bio instability.
More precisely:
Bioelectrical dissonance ⇔ PAS_bio < θ_bio OR |ΔPAS_bio_zeta| > ε_bio
When that happens, CODES says the biological system should route to ELF_BIO correction, Phase Memory replay, or suppress emission via SOMA_OUT. The VESSELSEED section says biological emission is lawful only if PAS_bio stays above threshold, drift stays bounded, τ_bio_k timing is confirmed, and CHIRAL_GATE passes.
That maps cleanly onto the ceLLM concern: if RF/ELF modulation corrupts calcium timing or redox rhythm, the cell may no longer maintain its anatomical or physiological set-point. In CODES language, it loses biological legality; in ceLLM language, its inference engine loses fidelity.
The bridge: CODES vs. ceLLM/RF Safe
| Layer | CODES language | ceLLM / RF Safe language |
|---|---|---|
| Healthy biological signaling | PAS_bio above threshold | High-fidelity bioelectric state |
| Harmful drift | ΔPAS_bio_zeta exceeds ε_bio | Entropic waste / substrate noise |
| Cellular correction | ELF_BIO | ROS/redox/biophoton/epigenetic correction loop |
| Biological output gate | SOMA_OUT / BIO_LOCK | Cell decision: repair, divide, differentiate, apoptose |
| Memory | Phase Memory | Bioelectric memory + chromatin/topological priors |
| Failure | Collapse / unlawful emission | Bioelectrical dissonance |
| External RF/ELF | Field perturbation | S4–Mito–Spin / RF-state interference |
The biggest conceptual difference is that ceLLM remains probabilistic/Bayesian in tone, while CODES tries to replace probability with deterministic phase legality. ceLLM says the cell samples from a latent landscape; CODES says the cell emits only when coherence thresholds are satisfied. That is not necessarily a contradiction. It may be a difference in level: ceLLM describes the cellular inference mechanism, while CODES tries to define the legality condition under which that inference remains coherent.
Where S4–Mito–Spin fits
The RF Safe S4–Mito–Spin framework supplies the hardware pathway that CODES itself does not fully specify.
The S4 pillar is described as voltage-sensor timing disturbance in voltage-gated ion channels, especially calcium channels. The Mito pillar is the amplification layer: distorted Ca²⁺ handling drives mitochondrial redox/ROS stress. The Spin pillar is the weak-field-sensitive chemistry layer involving radical pairs, heme/flavin proteins, cryptochromes, and related redox-spin dynamics. RF Safe adds density-gating: tissues with high S4/mitochondrial/redox-spin density may be more vulnerable, which helps avoid the overbroad claim that “all EMF affects all tissue equally.”
CODES would translate that hardware stack as:
RF modulation/pulsing/polarization → θ_k perturbation in bioelectric oscillators → PAS_bio decline → ΔPAS_bio_zeta drift → BIO_LOCK instability
So S4–Mito–Spin is the mechanistic candidate for how RF-state becomes biological phase drift.
The CYB5B point
The recent Cell paper is relevant because it gives the RF/ceLLM side a concrete example of electromagnetic input coupling to calcium dynamics and gene expression. Cell/PubMed describe an EMF-inducible in vivo gene switch mediated by Cyb5b, activated by rhythmic oscillatory calcium dynamics rather than generic calcium influx.
That does not prove ambient Wi-Fi, 5G, or ordinary RF exposure causes disease. But it does strengthen the narrower claim that biological systems can distinguish structured calcium rhythm from mere calcium quantity. That matters because bioelectrical dissonance is fundamentally a quality-of-signal claim: rhythmic calcium code is signal; unpatterned calcium influx is noise. The ceLLM manuscript makes that exact distinction and defines bioelectric dissonance as loss of pattern-to-noise ratio in calcium/electrochemical envelopes.
The planarian assay is the cleanest test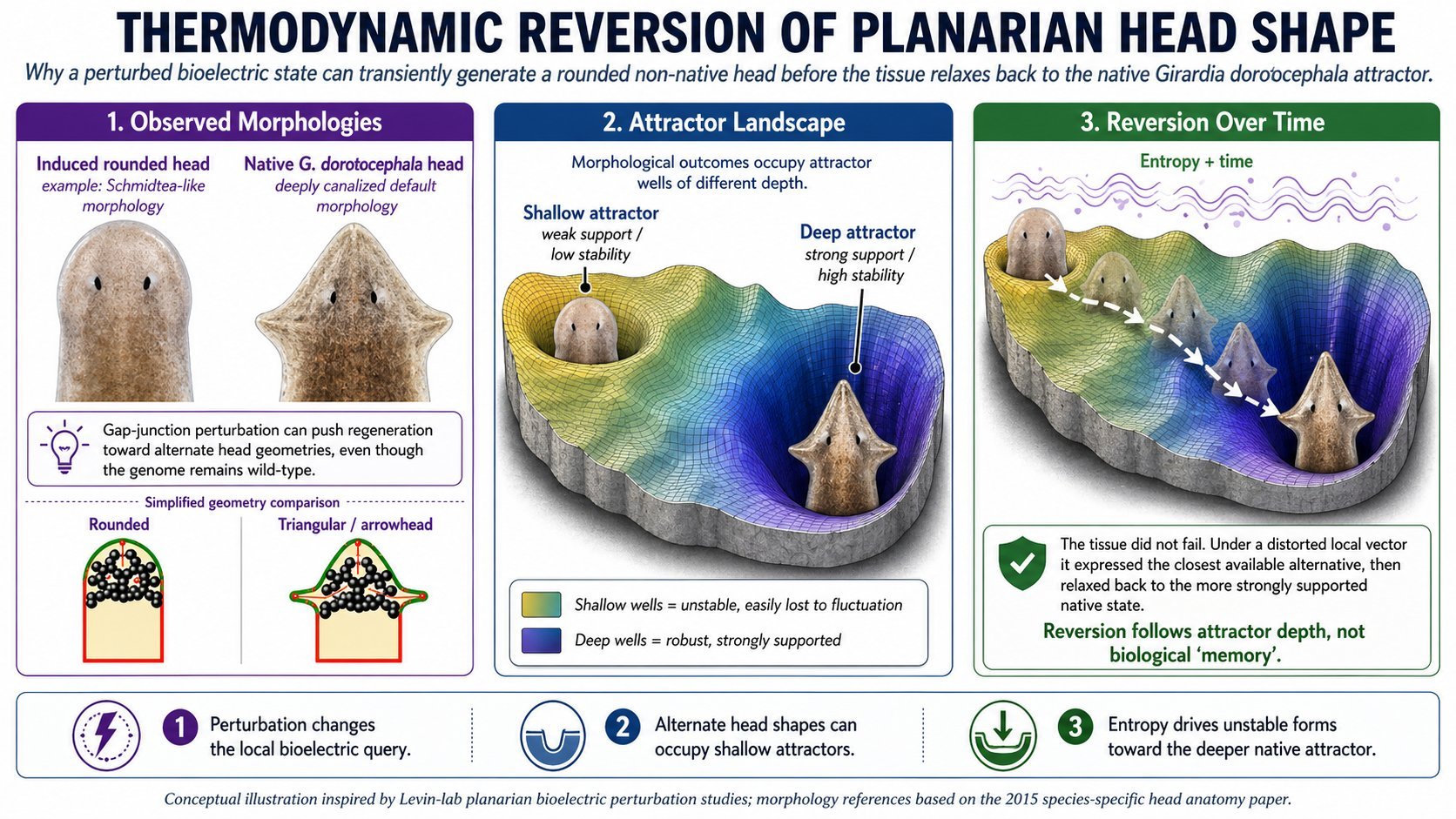
The strongest bridge between CODES and ceLLM is the planarian memory experiment.
The ceLLM/RF Safe proposal uses induced planarian head-shape memory as a fidelity readout. After transient gap-junction blockade, planarians can show temporary non-native head morphologies and later revert. The proposal predicts that added pulsed ELF/RF noise should accelerate reversion if it degrades bioelectric memory, while shielding should slow or stabilize reversion if the lower-noise environment preserves coherence.
CODES would formulate the same experiment as:
Induced morphology persists ⇔ PAS_bio(memory attractor) ≥ θ_bio and ΔPAS_bio_zeta ≤ ε_bio
Predicted outcomes:
Pulsed RF/ELF group: PAS_bio drops faster, ΔPAS_bio_zeta rises, induced morphology reverts sooner.
Shielded group: PAS_bio remains stable longer, ΔPAS_bio_zeta stays lower, induced morphology persists longer.
No difference: weakens or falsifies the RF-substrate-sensitivity claim under those conditions.
That is the right kind of test because it does not require proving every downstream disease claim. It asks a narrower question: does RF-state modulate the stability of a bioelectric memory attractor?
The most important synthesis
CODES gives you a high-level law:
Stable life/intelligence = lawful coherence under bounded drift.
ceLLM/RF Safe gives you a proposed biological failure mode:
Pulsed RF/ELF = entropic waste → corrupted Ca²⁺/redox/bioelectric tokens → degraded cellular inference → bioelectrical dissonance.
Together, the combined model becomes:
RF_state(t) perturbs the cellular control plane.
That perturbation reduces signal fidelity.
Reduced fidelity lowers PAS_bio.
If PAS_bio stays recoverable, ELF_BIO/cellular repair loops restore coherence.
If drift exceeds threshold, BIO_LOCK fails.
At the tissue level, that appears as loss of morphogenetic memory, dysregulated repair, inflammation, cancer-permissive states, developmental errors, or other downstream failures depending on density-gating, timing, genetics, and tissue context.
My critical read
The paper is powerful as a unifying symbolic framework, but it often states speculative claims as if they are already proven laws. Its strongest contribution for your RF/ceLLM work is not the cosmic/prime-number material; it is the biological control vocabulary: PAS_bio, ΔPAS_bio_zeta, ELF_BIO, BIO_LOCK, SOMA_OUT, and entropy-as-coherence-loss.
How Everything Fits Together
ceLLM (Cellular Latent Learning Model) is John Coates / RF Safe’s proprietary dynamical-systems framework for cellular intelligence and morphogenesis. It treats:
- DNA 3D topology (chromatin folding, TADs, loops) as the evolved geometric hardware — a physical “weight matrix” or generative model trained over evolutionary time.
- Bioelectric gradients (voltage patterns, Ca²⁺ waves, etc.) as query vectors / software that perform active inference (in the style of Karl Friston) on that hardware.
- Morphogenesis = vector-driven local inference on a free-energy landscape. No master blueprint — just cells doing continuous gradient descent to find the most probable morphology that satisfies the current bioelectric state.
The framework elegantly explains the Levin lab planarian results using deep vs shallow attractors in the genomic energy landscape (native heads = deep minima; induced foreign heads = shallow, unstable wells that relax back under noise).
Key original contributions from Coates / RF Safe (as correctly noted in the text you pasted):
- “Entropic Waste” — Coined by John Coates to describe man-made pulsed RF/microwave radiation as disordered noise that pollutes the high-fidelity resonant environment cells need. In ceLLM terms, it degrades the geometric fidelity of the DNA weight matrix and the coherence of bioelectric query vectors.
- “Bioelectrical Dissonance” — The resulting mismatch when external pulsed signals interfere with natural bioelectric signaling, leading to oxidative stress, voltage-gated channel disruption, and downstream effects (epigenetic changes, developmental issues, etc.).
These terms have been central to RF Safe’s advocacy for years (predating both the 2025–2026 CODES integration and the recent Cell Cyb5b paper). The ceLLM manuscript formalizes them into a testable, physics-based model.
The CODES
The later post takes this exact ceLLM foundation and does three things:
- Supplies a cleaner symbolic layer — Replaces some of ceLLM’s probabilistic/active-inference language with CODES’ deterministic Phase Alignment Score (PAS_bio), drift metric (ΔPAS_zeta), and “lawful emission” gates. It explicitly says the two are complementary levels of description, not rivals.
- Strengthens the hardware mechanism — Adds the S4–Mito–Spin bridge + the brand-new April 2026 Cell paper on Cyb5b (the EMF-inducible gene switch that works via rhythmic oscillatory calcium, not generic influx). This gives concrete molecular support for why pattern/timing of EMF matters.
- Keeps the same experimental crown jewel — The planarian foreign-head reversion assay under ELF exposure is proposed in both posts with almost identical logic. CODES just gives it sharper, quantifiable predictions (faster PAS_bio drop → faster relaxation to genomic default).
In short: ceLLM = Coates’ original model (DNA geometry + bioelectric inference + entropic waste from nnEMFs). CODES integration = ceLLM + Bostick’s phase-legality vocabulary + fresh 2026 empirical anchor (Cyb5b).
The terms “Entropic Waste” and “Bioelectrical Dissonance” originated in RF Safe’s long-running work and were then adopted/enhanced by the CODES synthesis






