
From RF Timing Noise to NMDAR Rescue
Publisher Note
This paper is written as a hypothesis-driven RF Safe position paper. It does not claim that non-native electromagnetic fields are the sole cause of autism, nor does it provide medical advice. The claim is narrower and more testable: RF/ELF exposures should be investigated as chronic upstream timing-noise stressors that may interact with genetic, chemical, immune, folate, sleep, and mitochondrial vulnerabilities during sensitive windows of neurodevelopment.
Figure 1. RF Safe conceptual pathway overview.
The figure presents a proposed pathway linking pulsed RF/ELF signal structure, ion-channel timing, mitochondrial sensing, calcium waveform disruption, chromatin/kinase state, NMDAR-related signaling, radial-glia persistence, and downstream neurodevelopmental vulnerability. It is a mechanistic model, not a direct proof of causation.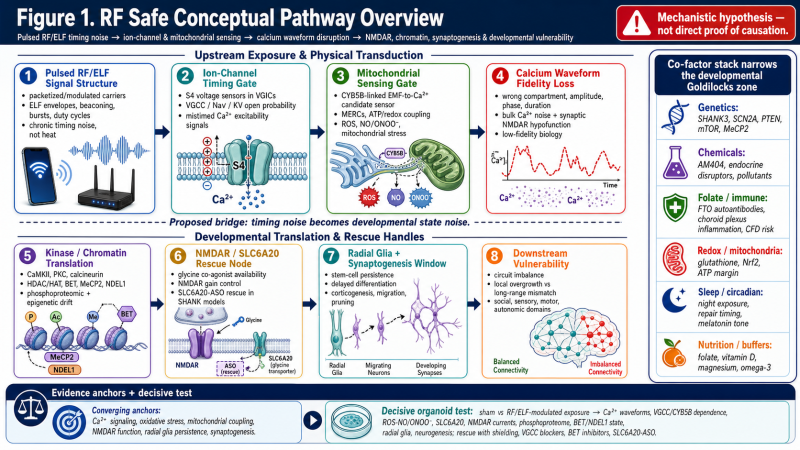
Abstract
A 2026 Nature Communications study reported that antisense oligonucleotide suppression of the glycine transporter Slc6a20a/SLC6A20 can restore NMDA receptor function in SHANK2– and SHANK3-mutant mouse models and in CRISPR-edited human cortical organoids. For RF Safe, the importance of this finding is not that it identifies the environmental origin of autism. It does not. Its importance is that it reveals an autism-relevant rescue node: glycine transport, NMDAR co-agonist availability, calcium-permeable synaptic signaling, and synaptic phosphoproteomic normalization.
This paper integrates that SLC6A20/NMDAR rescue finding with Martin Pall’s 2024 calcium-synaptogenesis model, Courchesne et al.’s 2024 embryonic brain cortical organoid findings, Cakir et al.’s 2025 RF/BET/radial-glia organoid findings, and Kim et al.’s 2026 CYB5B electromagnetic-field gene-switch paper. RF Safe’s synthesis is that non-native electromagnetic fields, or nnEMFs, should be investigated as a chronic upstream fidelity stressor. In this view, RF/ELF exposures do not need to be the sole cause of autism to matter. They need only lower the fidelity of ion-channel timing, mitochondrial calcium/redox signaling, kinase/phosphatase cycling, and NMDAR-linked synaptic development in children already carrying genetic or environmental vulnerabilities.
The practical conclusion is a research and precaution mandate: run mechanism-directed organoid, animal, and human epidemiology studies that measure calcium waveforms, CYB5B dependence, VGCC dependence, NMDAR function, SLC6A20 expression, NDEL1 activity, BET-mediated radial-glia differentiation, oxidative stress, folate biology, and phosphoproteomic state under realistic RF/ELF modulation conditions. Until such studies are done, RF Safe argues that reducing unnecessary RF exposure during pregnancy, infancy, sleep, and school is a rational low-regret public-health measure.
Executive Summary
The SLC6A20 paper is a downstream rescue paper. It shows that an autism-relevant signaling state can be pushed back toward normal by restoring glycine/NMDAR function and synaptic phosphorylation patterns.
Pall’s calcium model is an upstream convergence paper. It argues that EMFs, chemicals, and mutations can converge on intracellular calcium and six calcium-regulated mechanisms of synaptogenesis.
RF Safe’s contribution is the fidelity frame. The problem is not simply too much calcium or too little NMDAR signaling. It is calcium information appearing in the wrong compartment, wrong rhythm, wrong amplitude, and wrong developmental window.
The RF/BET organoid paper and the CYB5B gene-switch paper weaken the old assumption that non-thermal fields cannot couple to developmental biology. They do not settle autism causation, but they make the question biologically serious.
The research priority is no longer whether one exposure alone explains autism. The priority is whether chronic RF/ELF timing noise widens the effect of genetic, chemical, immune, folate, and mitochondrial risk factors during embryonic, fetal, and early-childhood windows.
1. The New SLC6A20 Paper Is a Rescue Map, Not an Origin Story
The 2026 SLC6A20 study should be read carefully. It does not claim that electromagnetic fields cause autism. It does not claim that one transporter explains the spectrum. What it does show is more precise and, for RF Safe, more important: an autism-relevant phenotype can sit in a persistent but modifiable signaling state.
In SHANK2- and SHANK3-mutant models, suppressed NMDAR function is not merely a passive consequence of a developmental lesion. It can be pharmacologically and molecularly re-tuned by suppressing Slc6a20a/SLC6A20, a transporter that helps regulate glycine availability. Glycine is a critical co-agonist for NMDAR activation. By increasing the local capacity for NMDAR co-activation in cognition-relevant regions, Slc6a20a/SLC6A20-ASO becomes a downstream handle on synaptic gain.
The decisive finding for this paper is phosphoproteomic. The authors report rescue of NMDAR hypofunction and synaptic phospho-proteomic profiles in prefrontal cortex, as well as rescue of suppressed NMDAR function in SHANK2- or SHANK3-mutant human cortical organoids. In other words, the therapeutic signal is not simply a behavioral effect. It is a correction of phosphorylation-state logic inside the synaptic network.
RF Safe interprets this as a downstream confirmation of the bioelectric-fidelity model. If correcting a co-agonist transporter can normalize NMDAR currents and synaptic phosphorylation profiles, then autism-relevant biology is not only a matter of fixed anatomy. It can also be a state problem: a network held in the wrong electrophysiological, metabolic, and phosphorylation regime.
2. Pall’s Model Gives the Calcium Convergence Layer
Martin Pall’s 2024 Brain Sciences paper offers a broad convergence model: during the perinatal period, excessive or otherwise inappropriate intracellular calcium can disrupt six mechanisms that control synaptogenesis: neuronal migration, dendritic outgrowth, synapse formation, synapse maturation, synaptic pruning, and MeCP2-linked epigenetic function.
Pall’s model places electronically generated EMFs upstream through voltage-gated calcium-channel activation and places many chemical classes upstream through NMDA receptor activation or other calcium-elevating mechanisms. The central idea is not that every autism case has the same cause. The central idea is that many causes can converge on calcium-controlled synaptogenesis.
RF Safe accepts the convergence layer but refines the language. The most defensible formulation is not simply that calcium is high. The stronger formulation is that calcium timing is low-fidelity. Calcium can be too high globally, too low synaptically, noisy in the wrong compartment, phase-shifted relative to gene-expression cycles, or prolonged after the developmental signal should have terminated. These are different failures, but they all damage biological computation.
That refinement resolves the apparent conflict between Pall’s excessive-calcium model and the SLC6A20 paper’s NMDAR hypofunction. A neuron can experience oxidative bulk calcium stress while simultaneously lacking clean synaptic NMDAR calcium transients. One is a noise burden; the other is a missing coincidence signal. Autism biology can plausibly involve both.
3. Calcium Is Not Just a Mineral; It Is a Timing Language
Cells compute with charge. Membrane potential, gap-junction coupling, sodium-channel excitability, calcium waves, mitochondrial redox state, and kinase/phosphatase cycles form a distributed information system. In the developing brain, that system controls cell division, migration, neurite extension, synapse formation, pruning, and activity-dependent gene expression.
This is why RF Safe uses the phrase low-fidelity biology. A low-fidelity system may still run, but its timing margins are degraded. A growth cone may still move, but the gradient it reads is noisy. A radial glial cell may still divide, but the decision to exit the stem-like state may be delayed. A synapse may still form, but NMDAR-dependent plasticity may misread coincidence. A kinase may still phosphorylate, but it may do so at the wrong time or on the wrong targets.
This is the bridge between EMF exposure, Tylenol/AM404, folate biology, NMDAR hypofunction, and autism genetics. They are not the same stressor. They are different levers on the same bioelectric code. Acetaminophen’s AM404 metabolite has now been shown to directly inhibit nociceptive voltage-gated sodium channels; SCN2A, encoding NaV1.2, is one of the best-known sodium-channel autism genes; and SLC6A20-ASO demonstrates that glycine/NMDAR co-agonism is a modifiable node in SHANK-related autism models. The shared substrate is ion-channel and synaptic timing.
RF Safe’s position is that a chronic field-level lever deserves the same seriousness as an episodic chemical lever. A drug exposure may be brief. A pulsed RF environment can be continuous: bedroom, router, phone, tablet, wearable, school network, smart meter, vehicle, tower, and neighbor devices. The biological question is whether that chronicity keeps vulnerable systems from returning to a clean baseline.
4. The RF Safe Extension: nnEMFs as Upstream Timing Noise
RF Safe does not argue that ambient wireless exposure is the only cause of autism. That claim would be too simple for a condition as heterogeneous as ASD. The position is more specific: nnEMFs may act as a baseline disruptor that reduces the fidelity of cellular signaling, making other insults more consequential and making some genetic vulnerabilities more penetrant.
In the RF Safe framework, the exposure is not treated as heat. It is treated as patterned physical information. Modern RF systems are not smooth biological silence. They contain modulation, duty cycles, packet structure, beaconing, burst timing, and low-frequency envelopes imposed on higher-frequency carriers. The concern is not that every signal is strong enough to cook tissue. The concern is that weak, repeated, polarized, and pulsed signals may bias probabilistic gates that biology uses for timing.
The first candidate gate is the S4 voltage sensor. Voltage-gated ion channels contain charged S4 helices that move in response to local electric fields. If an exogenous field biases open probability, even slightly, the effect can be amplified through calcium influx, neurotransmitter release, mitochondrial uptake, ROS production, and kinase activation.
The second candidate gate is mitochondrial. The 2026 CYB5B gene-switch paper reports an electromagnetic-field-inducible gene switch in which Cyb5b was identified as an essential mediator likely acting as an EMF sensor, with activation depending on rhythmic oscillatory calcium dynamics rather than generic calcium influx. RF Safe does not treat this as proof that Wi-Fi causes autism. RF Safe treats it as a proof-of-principle that biological systems can couple electromagnetic input to calcium-coded gene regulation through identifiable proteins.
Together, the S4 and CYB5B concepts form the physical entry layer of the model: one plasma-membrane route, one mitochondrial/redox route, both feeding the same calcium-phosphorylation-development axis highlighted by Pall and by the SLC6A20 rescue paper.
5. Embryonic Organoids Move the Timing Window Earlier
Courchesne and colleagues moved autism severity biology into the embryonic modeling window. Their 2024 Molecular Autism study generated thousands of brain cortical organoids from toddlers with ASD and controls. ASD organoids were enlarged, growth rate was accelerated, the largest organoids were linked to more severe social-symptom phenotypes, and NDEL1 activity/expression correlated with organoid growth rate and size.
This matters because it narrows the developmental search. If profound-autism biology can be visible in embryonic-stage cortical growth, then a major part of the causal architecture must operate before postnatal diagnosis, before school, before speech delay, and possibly before many parents know pregnancy has begun. That does not exclude later modifiers. It means early developmental timing is central.
Cakir and colleagues then supplied a directly relevant RF-organoid layer. In human cortical organoids exposed to RF between 800 and 2400 MHz, the authors reported altered radial glia differentiation, maintenance of stem-cell identity, delayed differentiation, induction of human endogenous retrovirus expression in neurons, and rescue of RF-induced developmental defects by BET inhibitors. This does not prove that everyday exposures reproduce the same outcome in vivo. It does show that RF can modulate human corticogenesis-relevant organoid biology through BET-mediated pathways.
RF Safe therefore sees three organoid layers converging. Courchesne shows early ASD severity can map to embryonic cortical growth and NDEL1. Cakir shows RF can alter radial-glia differentiation and BET-mediated pathways in human cortical organoids. Roh and colleagues show SLC6A20-ASO can rescue suppressed NMDAR function in SHANK2/SHANK3-mutant cortical organoids. These are not the same experiment. Their convergence is the point: early human cortical development is sensitive to growth, chromatin, radial-glia, NMDAR, and phosphorylation-state control.
6. NDEL1, BET Proteins, and SLC6A20 Are Not Separate Islands
NDEL1, BET proteins, SLC6A20, NMDARs, CaMKII, calcineurin, mitochondria, and VGCCs can be presented as separate mechanisms, but the cell does not experience them separately. They are coupled through timing.
NDEL1 is involved in proliferation, neurogenesis, neuronal migration, and cortical growth. BET proteins read acetylated chromatin and help decide whether progenitor programs stay open or close. SLC6A20 regulates a co-agonist environment that affects NMDAR current. NMDARs regulate calcium entry at synapses and downstream phosphorylation. CaMKII and phosphatases translate calcium timing into phosphorylation-state decisions. Mitochondria buffer calcium and convert timing failure into redox stress.
This is the key RF Safe bridge: a chronic upstream timing error does not need to directly mutate a gene to change developmental outcome. It can change how often gates open, how mitochondria decode calcium, how kinases phosphorylate targets, how chromatin is read, and how progenitors decide whether to divide, differentiate, migrate, or prune.
That is why the SLC6A20 rescue paper matters so much. It confirms that at least some ASD-relevant phenotypes can be shifted by retuning one transport/co-agonist gate. If one downstream gate can rescue NMDAR and phosphoproteome state, then the upstream question becomes urgent: what chronic environmental signals are pushing vulnerable systems into that state in the first place?
7. Tylenol, Folate, and Co-Factor Stacking
RF Safe’s model does not require denying chemical risk. It predicts chemical synergy. Acetaminophen, folate insufficiency, folate receptor alpha autoantibodies, maternal inflammation, endocrine disruptors, sleep disruption, nutrient deficiencies, and genetic risk variants can all narrow the system’s operating bandwidth.
The AM404 sodium-channel result is useful because it gives the public a simple analogy: a common exposure can directly modify ion-channel behavior. That does not mean acetaminophen is the sole cause of autism, and pregnant women should not make medication decisions without medical guidance. Fever and untreated pain can also carry risks. The point is mechanistic: ion channels are legitimate biological targets, and disturbances in their timing can matter during development.
Folate biology adds another layer. Neural-tube closure and early brain development require adequate methylation and nucleotide supply. A child can have adequate peripheral folate but inadequate CNS folate transport if folate receptor alpha autoantibodies or choroid plexus dysfunction are present. RF Safe proposes that mitochondrial/redox stress at barrier tissues could worsen this vulnerability, but this remains a hypothesis requiring direct testing.
The correct model is therefore not EMF versus Tylenol versus folate versus genetics. The correct model is bandwidth. The developing brain has a limited Goldilocks zone for bioelectric, metabolic, immune, and epigenetic fidelity. Multiple stressors can narrow that zone. RF Safe’s concern is that nnEMFs are continuous, ambient, and biologically plausible timing noise layered on top of all other stressors.
8. Why the Evidence Gap Is Not Reassurance
A fair critic can say that definitive human evidence connecting ambient RF exposure to autism is not established. RF Safe agrees with that narrow statement. But absence of definitive evidence is not evidence of absence when the decisive studies have not been funded or designed.
Most human RF health research has focused on cancer, adult exposure, phone-use proxies, or thermal safety assumptions. Very little has been built around embryonic cortical organoids, NMDAR electrophysiology, calcium waveform structure, CYB5B dependence, RF modulation patterns, maternal-fetal exposure reconstruction, or neurodevelopmental endpoints such as synaptogenesis and radial-glia timing.
The scientific question is now specific enough to test. Does RF/ELF exposure alter calcium waveforms in human cortical organoids? Does it shift NMDAR currents, NMDA/AMPA ratio, SLC6A20 expression, extracellular glycine, phosphoproteomic state, NDEL1 activity, or BET-dependent radial-glia differentiation? Are these effects blocked by shielding, VGCC blockers, CYB5B knockdown, antioxidant rescue, or SLC6A20-ASO? If the answer is no, the RF Safe model weakens. If the answer is yes, the regulatory paradigm changes.
9. The Decisive Experiment
The next study should not be another vague exposure study. It should be a mechanism-directed developmental-fidelity study.
RF Safe proposes human cortical organoids and, where appropriate, neural-tube and choroid-plexus organoid systems. Study groups should include control organoids, SHANK2/SHANK3-mutant organoids, idiopathic ASD-derived organoids, and organoids showing enlarged/profound-autism-like growth patterns. Exposure groups should include sham, RF carrier with minimal modulation, RF with realistic packetized modulation, ELF-only conditions, and combined chemical co-stressors under strict non-thermal monitoring.
Primary endpoints should include real-time calcium waveform metrics, VGCC dependence, CYB5B dependence, mitochondrial membrane potential, ROS/NO/ONOO⁻ markers, NMDAR mEPSCs, NMDA/AMPA ratio, extracellular glycine/proline/D-serine, SLC6A20 expression, phosphoproteomics, NDEL1 expression/activity, BET/chromatin marks, radial glia persistence, neurogenesis rate, neurite outgrowth, synapse formation, synapse maturation, and pruning markers.
The most important rescue arms would be shielding, VGCC blockers, CYB5B knockdown/rescue, antioxidant/Nrf2 support, BET inhibition, folate/folinic-acid support in appropriate models, and SLC6A20-ASO. The decisive prediction is simple: if nnEMF acts as upstream timing noise, then RF/ELF-exposed systems should show calcium waveform distortion and phosphoproteomic drift before gross malformation appears. If part of the drift runs through NMDAR co-agonist gating, then SLC6A20-ASO should rescue at least the NMDAR electrophysiology even if it does not rescue every developmental endpoint.
10. Public-Health Position
RF Safe’s public-health position follows from the mismatch between plausible biological risk and outdated regulatory framing. Current RF policy remains centered on thermal injury, yet the emerging mechanistic literature is increasingly about timing, modulation, calcium, mitochondria, redox, transcription, and developmental windows.
A precautionary response does not require fear or retreat from technology. It requires better engineering. Wired connections, fiber, low-emission device design, airplane mode during sleep, distance from the pregnant abdomen, reduced near-body use by children, and optical indoor communication are not radical measures. They are basic noise reduction.
Schools, nurseries, maternity environments, bedrooms, neonatal units, and fertility clinics should be treated as low-noise biological environments. RF Safe’s position is that society can keep connectivity while lowering unnecessary RF duty cycle in the places where development, sleep, repair, and reproduction occur.
Medical care remains medical care. Families should discuss medication, fever, folate, folinic acid, vitamin D, magnesium, mitochondrial support, and autoimmune testing with qualified clinicians. RF Safe’s role is not to replace medicine. It is to point out that the electromagnetic environment has become a continuous biological variable and must be studied and managed as one.
Conclusion
The SLC6A20-ASO study does not prove the RF Safe model. It does something more useful: it identifies a downstream rescue handle inside an autism-relevant synaptic state. Pall’s model identifies calcium-controlled synaptogenesis as a convergence layer. Courchesne identifies embryonic cortical overgrowth and NDEL1 as early severity-linked biology. Cakir identifies RF-sensitive BET/radial-glia effects in human cortical organoids. Kim identifies CYB5B-linked EMF-to-calcium gene-switch biology. Maatuf identifies a common drug metabolite that directly modulates voltage-gated sodium channels. SCN2A genetics confirms that ion-channel fidelity is already part of autism biology.
Taken together, these findings point to a coherent research agenda. Autism is not one thing, and RF is not a magic-bullet explanation. But non-native electromagnetic fields may be a chronic upstream timing-noise source acting on the same ion-channel, mitochondrial, calcium, redox, chromatin, NMDAR, and phosphorylation systems that modern autism biology keeps rediscovering.
RF Safe’s conclusion is therefore precise: the question is no longer whether a single exposure explains autism. The question is whether the modern RF/ELF environment lowers the fidelity of developmental signaling enough to raise the probability, severity, or persistence of atypical neurodevelopmental outcomes in vulnerable children. That question is testable. It is urgent. And until it is answered, lowering the noise floor around pregnancy and childhood is the rational path.
References
- Pall ML. Central Causation of Autism/ASDs via Excessive [Ca²⁺]i Impacting Six Mechanisms Controlling Synaptogenesis during the Perinatal Period: The Role of Electromagnetic Fields and Chemicals and the NO/ONOO⁻ Cycle, as Well as Specific Mutations. Brain Sciences. 2024;14(5):454. doi:10.3390/brainsci14050454.
- Roh JD, Bae M, Oh Y, et al. Glycine-modulating Slc6a20a-ASO restores NMDA receptor function in SHANK2 and SHANK3-mutant mice and cortical organoids. Nature Communications. 2026. doi:10.1038/s41467-026-73881-9.
- Courchesne E, Taluja V, Nazari S, et al. Embryonic origin of two ASD subtypes of social symptom severity: the larger the brain cortical organoid size, the more severe the social symptoms. Molecular Autism. 2024;15:22. doi:10.1186/s13229-024-00602-8.
- Cakir B, Tanaka Y, Choe MS, et al. Radiofrequency regulates the BET-mediated pathways in radial glia differentiation in human cortical development. Cell Reports. 2025;44(10):116238. doi:10.1016/j.celrep.2025.116238.
- Kim J, Hwang Y, Kim S, et al. Electromagnetic field-inducible in vivo gene switch for remote spatiotemporal control of gene expression. Cell. 2026. doi:10.1016/j.cell.2026.03.029.
- Maatuf Y, Kushnir Y, Nemirovski A, et al. The analgesic paracetamol metabolite AM404 acts peripherally to directly inhibit sodium channels. Proceedings of the National Academy of Sciences USA. 2025;122(23):e2413811122. doi:10.1073/pnas.2413811122.
- Kruth KA, Grisolano TM, Ahern CA, Williams AJ. SCN2A channelopathies in the autism spectrum of neuropsychiatric disorders: a role for pluripotent stem cells? Molecular Autism. 2020;11:23. doi:10.1186/s13229-020-00330-9.
- Shaw KA, et al. Prevalence and Early Identification of Autism Spectrum Disorder Among Children Aged 4 and 8 Years — Autism and Developmental Disabilities Monitoring Network, 16 Sites, United States, 2022. MMWR Surveillance Summaries. 2025;74(2).
- RF Safe. Neural Tubes, Autism, and Angel’s Fate on the 28th Day of Life: A Root-Cause Hypothesis for Non-Native EMFs as Entropic Waste. Internal RF Safe draft material, 2026.
- RF Safe. Why the EMF Timeline Aligns with Autism — And How Tylenol’s New Mechanism Helps Explain Why. Internal RF Safe draft material, 2026.






