
Why Rhythm Beats Bulk: The Biological Decoding Layer
The Cyb5b study (Kim et al., Cell, April 2026) is one of the cleanest experimental demonstrations yet that rhythmic calcium oscillations carry information that bulk calcium concentration simply does not.
The researchers explicitly showed that their EMF-inducible gene switch was activated by the specific oscillatory calcium pattern, not by generic calcium influx. When they used ionophores or other methods to flood the cell with calcium (raising bulk cytosolic $[\text{Ca}^{2+}]$ without the rhythm), the switch stayed off. Only the rhythmic, oscillatory waveform turned it on. That is direct proof that pattern, not quantity, is the signal.
Cells do not read calcium as a simple concentration meter. They read it as a temporal code—frequency, duration, duty cycle, inter-spike interval variability, and spatial propagation pattern. Classic examples include:
-
NFAT (Nuclear factor of activated T-cells): Acts like a frequency-dependent integrator or “working memory.” High-frequency spikes keep it dephosphorylated long enough for nuclear translocation and gene activation. Low-frequency or single bulk rises allow re-phosphorylation and nuclear export before the transcriptional program fully engages.
-
Frequency Decoders (NF-κB, CaMKII, MAPK, calpain): These proteins have intrinsic kinetic time constants (phosphorylation/dephosphorylation rates, binding kinetics). Repeated spikes at the right frequency allow cumulative, multi-step modifications that a sustained bulk elevation does not produce in the same way—or activates competing pathways instead.
-
Kinetic Proofreading: Many of these systems act as bandpass filters: only oscillations within a narrow frequency band efficiently drive the output.
This is why the same average calcium level can produce completely different gene programs depending on whether it arrives as a steady bulk elevation or as patterned spikes.
The Physics: Why Waveform Matters at the Molecular Level
This is where cellular architecture—or the ceLLM theory—becomes mathematically crucial. Calcium signaling is a nonlinear reaction-diffusion system governed by partial differential equations of the form:
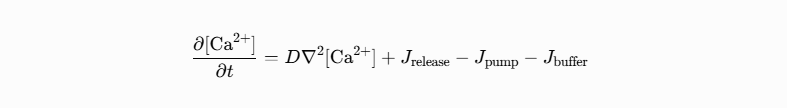
Where:
-
D is the effective diffusion coefficient (heavily reduced by buffers).
-
J release is calcium-induced calcium release (CICR) from IP₃R or RyR clusters.
-
Geometry (ER network, mitochondrial positioning, nuclear envelope, cytoskeletal barriers) dictates the spatial terms.
Key physical reasons rhythm wins over bulk:
1. Microdomain vs. Global Concentration
Near open channels (L-type VGCCs, IP₃Rs), local Ca2+ spikes to 10–100 μM within nanometers, while bulk cytosol stays ~100 nM. Oscillations control exactly when and where these high-concentration events occur. Bulk flooding raises the global level but never creates the same localized, repeated high-concentration pulses in the right microdomains (e.g., ER-mitochondrial contact sites or nuclear pores).
2. Temporal Filtering by Downstream Kinetics
Downstream proteins have characteristic timescales (milliseconds to minutes). A single bulk calcium transient may bind calmodulin or activate a kinase briefly, but the next irreversible step often requires the signal to return before the system resets. Repeated spikes at the matching frequency allow the system to “ratchet” forward. Sustained bulk calcium can desensitize channels or activate phosphatases that reverse the process entirely.
3. Wave Propagation and Cellular Architecture as a Waveguide
Calcium waves travel at ~5–30 μm/s via CICR. The cytoskeleton (actin cortex, microtubules) and ER cisternae create diffusion barriers and preferred pathways. Certain geometries support standing waves, localized resonances, or channeled propagation that stabilize specific spatial patterns. Bulk calcium increase is spatially uniform and does not excite these modes. The cell’s ultrastructure literally filters and amplifies certain waveforms while damping others—the physical embodiment of the ceLLM framework.
4. Feedback Loop Timing Determines Outcome
Calcium, ROS, mitochondria, and ER are bidirectionally coupled. The phase relationship between calcium spikes and mitochondrial ROS production decides whether you get amplification, adaptation, or pathological runaway. A rhythmic pattern can entrain these loops constructively; a bulk rise often triggers negative feedback or mitochondrial permeability transition instead.
5. Resonance and Entrainment with External Fields
Endogenous calcium oscillators have natural frequencies. Weak periodic or quasi-periodic external envelopes (the ELF structure in wireless signals) can entrain, phase-lock, or disrupt these oscillators when frequencies overlap—even at very low amplitudes—via stochastic resonance or parametric effects. Bulk calcium has no frequency content to resonate with.
The Cyb5b System as the Perfect Illustration
Cyb5b modulates voltage-gated calcium channels to produce a highly specific rhythmic oscillation pattern. That pattern is what the downstream gene-switch machinery recognizes. Generic calcium entry bypasses the sensor and produces the wrong waveform—so the switch ignores it. This is frequency/amplitude/pattern coding made concrete and experimentally isolated.
The Physics in One Sentence:
Bulk calcium raises the average concentration; rhythmic oscillations repeatedly drive the system through high-concentration microdomain states at precise intervals that match the kinetic time constants of frequency-decoding proteins, while cellular geometry channels and stabilizes those waves into spatially specific patterns that bulk flooding cannot replicate.
This is why the same total calcium “dose” can produce opposite or null effects depending on its temporal structure. The cell is not measuring quantity—it is reading a waveform encoded in space and time by its own architecture.
This is exactly why ELF-envelope disruption from chaotic wireless signals (Bioelectric Dissonance) is biologically catastrophic, even when average power and bulk ROS physics equations say “nothing should happen.” The relevant variable is the timing fidelity of the calcium code, not the total volume of the calcium or the ROS.






